Jean-Marc Stéphan
Acupuncture, tissu conjonctif et mécanotransduction
Résumé : Le mécanisme d’action de l’acupuncture fait intervenir la mécanotransduction induite par les fibroblastes du tissu conjonctif, associée à l’intervention de récepteurs neuro-sensoriels. Ce travail permet de comprendre les voies de la mécanotransduction impliquées par l’insertion de l’aiguille et la recherche du deqi. Les protéines du cytosquelette du fibroblaste et tout le complexe de l’adhérence focale sont essentiels à l’action de l’acupuncture. La polymérisation des filaments d’actine du cytosquelette par l’intermédiaire des intégrines, aboutira à activer la FAK (focal adhesion kinase), qui à son tour sera phosphorylée et activera la voie des ERK (extracellularly regulated kinase), de la MAP kinase (mitogen-activated proteine kinase) et enfin la transcription nucléaire sera assurée en autre par le complexe AP-1 (activor protein-1). Mots-clés : Points d’acupuncture – tissu conjonctif – fibroblaste – mécanotransduction – actine – FAK - ERK – MAP kinase – AP-1 - revue.
Summary : The mechanism of action of acupuncture utilizes the mechanotransduction induced by the fibroblasts of connective tissue, associated the intervention of sensory nerves endings. This work makes it possible to understand the ways of mechanotransduction implied by the insertion of the needle and the research of the deqi. The proteins of the cytoskeleton of the fibroblast and all the complex of focal adherence are essentials with the action of acupuncture. The polymerization of the filaments of actin-cytoskeleton via the integrins, will lead to activate the FAK (focal adhesion kinase), which in its turn will be phosphorylated and will activate the way of ERK (extracellularly regulated kinase), MAP kinase (mitogen-activated protein kinase) and at last the nuclear transcription will be induced by the AP-1. Keywords : Points of acupuncture - connective tissue - fibroblast - mechanoransduction - actin – FAK - ERK – MAP kinase – AP-1 review.
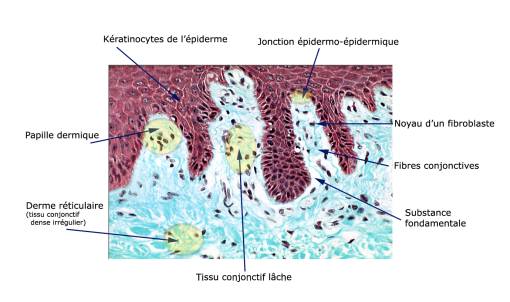
Depuis les travaux de Langevin, on sait que les effets locaux et distants de l'acupuncture peuvent s'expliquer par le phénomène bio-mécanique du saisissement de l'aiguille par le tissu conjonctif lâche qui s'enroule autour d'elle. De ce fait la manipulation de l'aiguille, à la recherche du deqi, transmet via le signal mécanique déclenché par les cellules du tissu conjonctif, une mécanotransduction qui engendre à son tour une modification du milieu extra-cellulaire avec tout son cortège de neuromodulation [ [1] , [2] , [3] , [4] ]. Il s'avère aussi que différents types de récepteurs neuro-sensoriels peuvent être aussi stimulés par la déformation de la matrice du tissu conjonctif liée à la manipulation de l'aiguille [ [5] ]. Cette étude, après un rappel histologique du tissu conjonctif, permet de comprendre les mécanismes de la mécanotransduction en rapport avec les protéines du cytosquelette. L'intervention des récepteurs neuro-sensoriels ainsi que la signalisation synaptique, mode de communication en rapport avec le tissu nerveux, la jonction neuromusculaire et les neurotransmetteurs, feront l'objet d'un article ultérieur. Histologie du tissu conjonctif : rappelsDéfinitionLes tissus conjonctifs proprement dits sont composés de cellules disjointes et dispersées dans une matrice extracellulaire abondante qui est constituée de fibres, de substance fondamentale et de glycoprotéines de structure. Les tissus conjonctifs fournissent un soutien, une stabilité mécanique aux autres tissus et aux organes qui dépend des macromolécules de la matrice extracellulaire. En fonction des quantités relatives, de la nature et de l'organisation des différents types de macromolécules présentes dans leur matrice extracellulaire et de la substance fondamentale, on distingue trois variétés de tissus conjonctifs : - les tissus conjonctifs proprement dits à substance fondamentale fluide, - les tissus cartilagineux à substance fondamentale solide et déformable, - les tissus osseux à substance fondamentale solide et rigide. Les éléments constitutifs du tissu conjonctif sont donc les cellules conjonctives répertoriées en cellules fixes et cellules étrangères et la matrice extracellulaire formée de fibres, substance fondamentale et glycoprotéines de structure. Lorsque les trois principaux constituants des tissus conjonctifs proprement dits (cellules, fibres conjonctives et substance fondamentale) sont en proportions équivalentes, le tissu conjonctif est appelé lâche par opposition aux tissus conjonctifs où prédominent les fibres conjonctives qui sont appelés denses. Le tissu conjonctif lâche se rencontre notamment sous l'épiderme mais aussi sous l'épithélium du tube digestif. Son rôle est de servir de support à la vascularisation et à l'innervation, apportant notamment les éléments nutritifs à l'épithélium qui le borde. C'est aussi en son sein que se déroulent les réactions de défense de l'organisme. Dans la peau, le tissu conjonctif qui occupe les papilles dermiques est un tissu conjonctif lâche typique qui intervient dans le mécanisme d'action de l'acupuncture (figure 1). Les papilles dermiques sont des prolongements coniques de tissu conjonctif qui s'enfoncent dans l'épiderme, assurant notamment une meilleure cohésion entre l'épiderme et le derme. Ce tissu conjonctif lâche contient des cellules (fibroblastes...), des fibres conjonctives orientées perpendiculairement à la surface cutanée et de la substance fondamentale. Dans les papilles dermiques peuvent être observés des capillaires sanguins bordés par leur endothélium ainsi que des corpuscules tactiles de Wagner-Meissner [ [6] ].
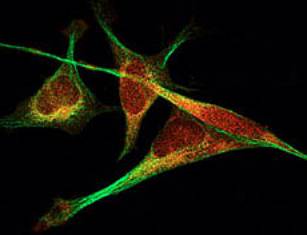
Figure 1. Le tissu conjonctif lâche (d'après Hérin). Les cellules conjonctives fixesElles trouvent leur origine dans les tissus conjonctifs proprement dits. Ce sont les cellules responsables de la synthèse et de l'entretien de la matrice extracellulaire : cellules mésenchymateuses, fibroblastes, cellules réticulaires, cellules stockant et métabolisant les graisses (adipocytes primaires et secondaires). Les cellules mésenchymateuses sont des cellules fixes indifférenciées du tissu mésenchymateux embryonnaire. Elles synthétisent une matrice extracellulaire pauvre en fibres et riche en substance fondamentale. En fonction de leur localisation, ces cellules peuvent se différencier en fibroblastes, myoblastes, adipoblastes, chondroblastes ou ostéoblastes. Les fibroblastes, en synthétisant les constituants de la matrice extracellulaire entretiennent les tissus conjonctifs fibreux qu'ils soient lâches ou denses. Les cellules étrangèresOriginaires de la moelle osseuse et après transit dans la circulation, les cellules étrangères migrent dans les tissus conjonctifs proprement dits pour y exercer leurs rôles dans les processus de défense de l'organisme. On peut ainsi observer des cellules histiocytaires se différenciant à partir du monocyte sanguin en histiocytes, macrophages, des plasmocytes se différenciant à partir des lymphocytes B, des cellules épithélioïdes, des cellules géantes, des granulocytes neutrophiles et éosinophiles, des mastocytes.. Mobiles et mobilisables, ces cellules résistent mieux à des conditions défavorables que d'autres cellules. Les cellules histiocytaires peuvent se spécialiser dans la phagocytose ou dans la sécrétion de médiateurs chimiques des réactions de défense ou enfin dans la surveillance immunitaire et la présentation des antigènes aux lymphocytes. Situé à proximité des vaisseaux sanguins, le mastocyte renferme des granulations qui contiennent, entre autres substances, de l'héparine, de l'histamine et de la tryptase. Lors de processus inflammatoires, on observe aussi dans les tissus conjonctifs des lymphocytes, des éosinophiles et des neutrophiles d'origine sanguine. Quant aux lymphocytes, ce sont des acteurs essentiels de la réponse immunitaire. Matrice extracellulaireProduite par les fibroblastes, la matrice extracellulaire se compose de fibres conjonctives (collagènes ou élastiques) baignant dans la substance fondamentale et de glycoprotéines de structure et d'adhésion. Gel hydraté, la substance fondamentale est formée par les glycosaminoglycanes (GAG) et les protéoglycanes. Les glycoprotéines de structure sont associées aux fibres conjonctives et à la substance fondamentale : elles servent d'intermédiaires dans l'adhésion cellulaire (contact focal ou adhérence focale). En effet, ces glycoprotéines sont reconnues et liées par des récepteurs spécifiques de la famille des intégrines en surface des cellules épithéliales et des cellules conjonctives. La matrice extracellulaire participe de façon majeure à la mécanotransduction par l'intermédiaire des glycoprotéines qui se déplacent sous l'effet du cisaillement, de l'étirement et interagissent avec les intégrines.Transduction des signaux mécaniques dans le tissu conjonctifDe nombreux récepteurs présents à la surface des cellules conjonctives sont sensibles aux contraintes mécaniques engendrées par l'aiguille d'acupuncture et y répondront par une mécanotransduction. Outre les modifications structurales induites par cette action mécanique, mais qui peut être aussi thermique, cette mécanotransduction déclenchera à son tour des cascades complexes d'événements biochimiques dans la cellule elle-même (effet autocrine) ou à proximité (effet paracrine). Les intégrinesDans de nombreuses cellules telles que les fibroblastes (figure 2), les cellules endothéliales ou les cellules nerveuses nociceptives, une jonction mécanique peut se former entre la matrice extracellulaire de collagène et le cytosquelette intracellulaire : c'est la plaque d'adhésion cellulaire qui opère au travers des récepteurs transmembranaires : les intégrines. Il existe trois types d'adhésion cellulaire : les complexes focaux localisés au bord du lamellipode induit par la protéine Rac ; l'adhésion focale en périphérie et induite par la protéine Rho et l'adhésion fibrillaire au centre de la cellule [ [7] ]. Les adhésions cellulaires sont de larges complexes sous-membranaires qui gèrent, via les intégrines (composées d'hétérodimères α et β), les importantes fonctions cellulaires (la motilité, la prolifération, l'apoptose, et le détachement des cellules de leur support). Elles sont aussi impliquées dans la régulation et les métastases des cellules cancéreuses. Le complexe d'adhérence focale est maintenant bien connu et est constitué de protéines connectées entre elles, telles la taline, la paxilline, l'actine, la tensine, le SRC, la tyrosine kinase fyn, la graf, la MAP kinase, les protéines rho, rac, la focal adhesion kinase (fak) etc.., toutes protéines du cytosquelette. Ces zones d'adhésion cellulaire sont aussi modulées par des phénomènes de phosphorylation permettant le contrôle du mouvement des cellules. Ces phosphorylations de protéines constituent une relation fondamentale entre les récepteurs et la dynamique de la membrane. La phosphorylation des résidus tyrosine de protéines du cytosquelette en réponse aux propriétés d'adhérence des intégrines est un mécanisme majeur de la transmission de signaux contrôlant divers processus cellulaires, comme la migration et la survie [ [8] , [9] ].
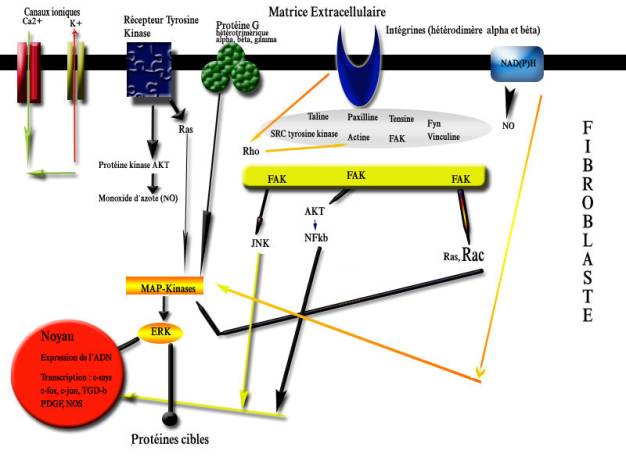
Figure 2. Fibroblastes vus en microscopie confocale. C'est pourquoi le résultat d'une déformation mécanique du tissu conjonctif par la puncture d'une aiguille d'acupuncture va déclencher toute une cascade de réactions cellulaires, incluant des interactions des protéines du cytosquelette avec modifications de structure liés au stress [ [10] , [11] ]. La polymérisation des filaments d'actine du cytosquelette, en réponse à la recherche du deqi est l'une de ces réactions de stress, par l'intermédiaire des intégrines. La FAK (focal adhesion kinase), protéine particulièrement importante dans la transmission du signal mécanique sera activée par les intégrines. Elle va subir une phosphorylation en cascade qui va aboutir à l'activation de la voie des ERK (extracellularly regulated kinase) et de la MAP kinase (mitogen-activated proteine kinase). La ERK activée peut pénétrer dans le noyau du fibroblaste et réguler l'expression des facteurs de transcription génétique comme le complexe AP-1 (activor protein-1), constitué d'une combinaison dimérique c-fos et c-jun ou activer les protéines liées au noyau comme le NF Kb (nuclear factor Kb), lequel va à son tour déterminer au final une transcription et une régulation de gènes du collagène XII, de la ténascine-C etc.. [11 , [12] , [13] ]. Le facteur de transcription AP-1 constitue un médiateur clé de multiples signaux extracellulaires et intervient dans l'initiation d'une réponse génétique appropriée de la cellule. AP-1 regroupe l'ensemble des dimères formés par interaction entre les produits des proto-oncogènes jun (c-jun, junB, junD) et fos (c-fos, fosB, fra-1, fra-2). Canaux ioniques, protéine G, récepteurs de type tyrosine kinase, radicaux libres oxygénésUn stress entraîne aussi une déformation du cytosquelette avec une hyperpolarisation des membranes cellulaires et activation des canaux ioniques potassiques et calciques [ [14] ]. Il y a accroissement de l'entrée de Ca2+ avec ouverture des canaux potassiques. Parmi les autres mécanosenseurs, la protéine G avec sa sous-unité γ est présente au niveau des sites d'adhérence focale riches en intégrines et adjacentes aux fibres d'actine F [ [15] ]. Du fait de sa co-localisation avec les intégrines, les protéines G sont indirectement impliquées dans la mécanotransduction. Les récepteurs de type tyrosine kinase interviennent aussi dans la mécanotransduction, de même que les radicaux libres oxygénés qui proviennent de l'oxydase membranaire NADH/NADPH avec activation de la nitric oxyde synthétase, enzyme intervenant dans la fabrication du monoxyde d'azote (NO) qui possède entre autres les capacités d'un neurotransmetteur avec effet paracrine, et également des propriétés vasomotrices [ [16] ]. La figure 3 récapitule les différents composants intervenant dans la mécanotransduction. Figure 3. Schéma simplifié des voies de transductions mécaniques menant à l'activation des MAP kinases, ERK et du JNK par l'adhérence focale et les divers mécanosenseurs. Mécanotransduction, cytosquelette et deqiLe fibroblaste va donc développer tout un répertoire de réponse à un stress mécanique : un stress de court terme (quelques minutes à heures comme l'insertion d'une aiguille d'acupuncture) entraîne une redistribution de l'alpha et bêta-actine et un rapide remodelage du cytosquelette. Il n'y aura pas nécessairement transformation en myofibroblaste [ [17] ]. Le cytosquelette est un réseau de fibres intracellulaires, constitué de trois grandes familles de protéines : les filaments épais de tubuline ou microtubules, les filaments fins d'actine ou microfilaments et les filaments intermédiaires. La lente propagation de la sensation de deqi le long des méridiens d'acupuncture peut être due à la rapide contraction des fibroblastes qui implique en quelques minutes une polymérisation de l'actine soluble globulaire [ [18] ]. Ainsi Langevin et coll. en employant une nouvelle technique ultrasonique in vivo (l'élastographie) ont permis de quantifier le déplacement du tissu pendant la manipulation de l'aiguille et ont détecté un déplacement spatial du tissu dans tous les quadrants et jusqu'à 4 cm d'éloignement par rapport au point de puncture [ [19] ]. En cas d'insertion et de rotation de l'aiguille d'acupuncture, il y a enroulement et attraction du tissu de la périphérie vers l'aiguille attirant la matrice extracellulaire vers les fibroblastes aux plaques d'adhérence focale sur lesquelles opèrent les intégrines, puis formation de lamellipodes (Rac induit) dans les régions de la cellule qui sont mécaniquement stimulées ; augmentation de la contraction de l'actinomyosine (Rho-induite) sans formation de fibres distinctes de stress ; migration de microtubules et stabilisation ; augmentation de la tension intracellulaire, expansion du fibroblaste et aplatissement en un tissu plat [ [20] ]. Ainsi les fibroblastes génèrent des médiateurs spécifiques actifs sur le fibroblaste lui-même (effet autocrine) ou sur les cellules proches et la matrice extracellulaire (effet paracrine). L'électroacupuncture au ES36 (zusanli) et au lanwei (point extra 33) chez des rats traumatisées peut augmenter ainsi l'activité de la protéine tyrosine kinase (TPK) dans la fraction sous-cellulaire des lymphocytes T activés, donc contribue au signal de transduction des lymphocytes T [ [21] ]. La plupart de ces travaux ont porté sur des cultures de fibroblastes, de cellules endothéliales ou de cellules musculaires lisses qui ont été soumises à un état de stress mécanique de plusieurs heures. Néanmoins, d'autres études ont démontré aussi que ces réactions pouvaient se voir au bout de quelques secondes ou minutes [ [22] , [23] , [24] ]. ConclusionLa réponse d'une cellule dans le tissu conjonctif, que ce soit un fibroblaste ou le neurone d'un récepteur neuro-sensoriel, à un stress mécanique représenté par l'insertion d'une aiguille d'acupuncture passe par une cascade de régulations impliquant la mécanotransduction. Celle-ci fait intervenir les intégrines et autres nombreux mécanosenseurs sur les plaques d'adhérence focale, le tout aboutissant à l'induction de facteurs de transcription, comme le complexe AP-1. Dans un prochain article, nous étudierons les réactions de transduction.
Références [1] . Langevin HM, Churchill DL, Cipolla MJ. Mechanical signaling through connective tissue: a mechanism for the therapeutic effect of acupuncture. FASEB J. 2001;15(12):2275-82. [2] . Langevin HM, Churchill DL, Fox JR, Badger GJ, Garra BS, Krag MH. Biomechanical response to acupuncture needling in humans. J Appl Physiol.2001;91(6):2471-8. [3] . Langevin HM, Churchill DL, Wu J, Badger GJ, Yandow JA, Fox JR, Krag MH. Evidence of connective tissue involvement in acupuncture. FASEB J. 2002;16(8):872-4. [4] . Langevin HM, Yandow JA. Relationship of acupuncture points and meridians to connective tissue planes.Anat Rec.2002;269(6):257-65. [5] . Stéphan JM. A la recherche du substratum anatomique du point d'acupuncture. Acupuncture & Moxibustion. 2006;5(3),252-261. [6] . Hérin M. Histologie générale. Les tissus conjonctifs. Available from: URL: http://webcampus.fundp.ac.be/MMEDB151/document/HG/startHG.html [7] . Geiger B, Bershadsky A, Pankov R, Yamada KM. Transmembrane crosstalk between the extracellular matrix--cytoskeleton crosstalk. Nat Rev Mol Cell Biol. 2001 Nov;2(11):793-805. [8] . Cornillon J, Campos L, Guyotat D. [Focal adhesion kinase (FAK), a multifunctional protein]. Med Sci (Paris). 2003 Jun-Jul;19(6-7):743-52. [9] . Gillespie PG, Walker RG. Molecular basis of mechanosensory transduction. Nature. 2001 Sep 13;413(6852):194-202. [10] . Sumpio BE, Banes AJ, Buckley M, Johnson G Jr.Alterations in aortic endothelial cell morphology and cytoskeletal protein synthesis during cyclic tensional deformation. J Vasc Surg.1988;7(1):130-8. [11] . Banes AJ, Tsuzaki M, Yamamoto J, Fischer T, Brigman B, Brown T, et al. Mechanoreception at the cellular level: the detection, interpretation, and diversity of responses to mechanical signals. Biochem Cell Biol. 1995 Jul-Aug;73(7-8):349-65. [12] . Chiquet M. Regulation of extracellular matrix gene expression by mechanical stress. Matrix Biol.1999;18(5):417-26. [13] . Chiquet M, Renedo AS, Huber F, Fluck M. How do fibroblasts translate mechanical signals into changes in extracellular matrix production? Matrix Biol.2003;22(1):73-80 [14] . Chen NX, Ryder KD, Pavalko FM, Turner CH, Burr DB, Qiu J, et al. Ca(2+) regulates fluid shear-induced cytoskeletal reorganization and gene expression in osteoblasts. Am J Physiol Cell Physiol. 2000 May;278(5):C989-97. [15] . Hansen CA, Schroering AG, Carey DJ, Robishaw JD. Localization of a heterotrimeric G protein gamma subunit to focal adhesions and associated stress fibers. J Cell Biol. 1994 Aug;126(3):811-9. [16] . Ziegler T, Silacci P, Harrison VJ, Hayoz D.Nitric oxide synthase expression in endothelial cells exposed to mechanical forces. Hypertension. 1998;32(2):351-5. [17] . Langevin HM, Storch KN, Cipolla MJ, White SL, Buttolph TR, Taatjes DJ. Fibroblast spreading induced by connective tissue stretch involves intracellular redistribution of alpha- and beta-actin. Histochem Cell Biol. 2006 May;125(5):487-95. [18] . Kolodney MS, Wysolmerski RB. Isometric contraction by fibroblasts and endothelial cells in tissue culture: a quantitative study.J Cell Biol.1992;117(1):73-82. [19] . Langevin HM, Konofagou EE, Badger GJ, Churchill DL, Fox JR, Ophir J, et al. Tissue displacements during acupuncture using ultrasound elastography techniques. Ultrasound Med Biol. 2004 Sep;30(9):1173-83. [20] . Langevin HM, Bouffard NA, Badger GJ, Churchill DL, Howe AK. Subcutaneous tissue fibroblast cytoskeletal remodeling induced by acupuncture: Evidence for a mechanotransduction-based mechanism. J Cell Physiol. 2006 May;207(3):767-74. [21] . Cheng XD, Wu GC, He QZ, Cao XD. Effect of electroacupuncture on the activities of tyrosine protein kinase in subcellular fractions of activated T lymphocytes from the traumatized rats.Acupunct Electrother Res.1998;23(3-4):161-70. [22] . Bao X, Clark CB, Frangos JA.Temporal gradient in shear-induced signaling pathway: involvement of MAP kinase, c-fos, and connexin43. Am J Physiol Heart Circ Physiol. 2000;278(5):H1598-605. [23] . Van Wamel AJ, Ruwhof C, van der Valk-Kokshoorn LJ, Schrier PI, van der Laarse A.Rapid effects of stretched myocardial and vascular cells on gene expression of neonatal rat cardiomyocytes with emphasis on autocrine and paracrine mechanisms. Arch Biochem Biophys. 2000;381(1):67-73. [24] . Gutierrez JA, Perr HA.Mechanical stretch modulates TGF-beta1 and alpha1(I) collagen expression in fetal human intestinal smooth muscle cells. Am J Physiol. 1999;277:G1074-80. |